Tu cesta
Tu cesta

Serotonin
26 Dec 2016
Serotonin es el biogenic amine que es formado del aminoácido tryptophan por su hydroxylation y decarboxylation. Una cantidad significativa de un serotonin contiene en las células enterokhromaffinnykh del intestino, CNS, principalmente en hypothalamus y mesencephalon, plaquetas, cantidad más pequeña en el labrotsitakh, células del mástil, cápsulas suprarrenales. Un serotonin ejerce el impacto a actividad nerviosa, reducción de causas de musculation liso de un intestino, útero, bronquios, y también estrechamiento de buques. Las reacciones de un organismo a un serotonin son la piedra angular el central, myotropic, ganglionic, efectos reflejos.
Educación. Un serotonin (5-gidroksi-triptamin, 5-HT) se sintetiza en las células del epitelio intestinas enterokhromaffinnykh de un L-tryptophan. Un serotonin también se forma en células nerviosas de mezenterialny plexus y en un CNS donde desempeña un papel del mediador. Las plaquetas no sintetizan un serotonin, sin embargo lo toman y se acumulan.
Receptores de Serotonin. Hay varios tipos de receptores. De los receptores del punto de vista farmacológicos 5-HT1 y 5-NT2, y también 5-HT3 y 5-HT4 son importantes. Por regla general, los receptores trabajan a través de la G-proteína. El receptor 5-HT3 representa el canal cationic.
Efecto de serotonin. Sistema cardiovascular. La influencia de serotonin en el sistema cardiovascular es compleja ya que el serotonin tiene el efecto (a veces enfrente mandado) con muchos sitios a través de tipos diferentes de receptores. A través de 5-HT2-retseptory localizado en músculos no estriados de paredes de buques, un serotonin causa el estrechamiento de buques. El efecto al mismo tiempo indirecto de serotonin puede llevar a vasodilatation y la depresión del ABP: a través de 5-HT1A-retseptory bloquea neuronas comprensivas de un brainstem y la periferia que lleva a la depresión de tonus comprensivo; la activación de receptores 5-HT1 en endothelium vascular lleva a la emisión de vazodilatator (prostatsickin, NO). El serotonin que se suelta de plaquetas desempeña un papel en procesos de thrombogenesis, hemostasis y en hypertonia pathogenesis en mujeres embarazadas.
Sumatriptan (un agente contra la migraña) es un 5-HT1D agonist y 5-HT1B-retseptorov. El estrechamiento de buques craneales causados por el bloqueo de la emisión de neuropeptids lleva a un llamado �neurogenic inflamación� o dirigir la crisis vascular. Al mismo tiempo puede haber un sentimiento de coacción en un pecho ligado a una restricción espástica de arterias coronarias. Otros �tryptones� son naratriptan, zolmitriptan y rizatripan.
Aparato digestivo. El serotonin formado en mezenterialny plexus o células enterokhromaffinnykh estimula un peristalsis y la secreción de un intestino por medio de la influencia en 5-HT4-retseptory.
Las tentativas del impacto farmacológico a un motility de un intestino a través de receptores serotonin eran ineficaces hasta ahora. Tsizaprid, un 5-HT4 agonist, sin embargo, puede aumentar motility propulsivo de un intestino. Este efecto secundario es bloqueado por Atropinum que demuestra la rebelión de la concentración de Acetylcholinum bajo la influencia de tsizaprid. La medicina da un juego de efectos secundarios. Es inactivated en el mecanismo con la participación de SURZA4 que lleva a varias interacciones imprevistas con otras sustancias. Tsizaprid causa perturbaciones (incluso el serio) un ritmo cordial (la reducción de intervalos QT) por lo tanto la medicina no encontró la aplicación.
Sistema nervioso central. Los receptores de Serotonin desempeñan un papel importante en varias funciones CNS.
Fluoxetine es el antidepresivo que bloquea la vuelta serotonin nuevo consumo. Bastante aumenta la motivación, y también baja el apetito.
Sibutramin, blocker de la vuelta nuevo consumo serotonin neuronal y Noradrenalinum; se aplica como el remedio por la obesidad.
Ondansetron tiene el efecto antivomitivo expresado en los vómitos causados por la aplicación de tsitostatik. Es un antagonista de 5-HT3-retseptorov. Los análogos de ondansetron son tropisetron y granisetron.
El ácido y los otros de psychedelia (psikhotomimetika), como la mescalina y psilocybin, causan alucinaciones, desórdenes de conocimiento, miedo, quizás, debido a la activación de 5-NT-retseptorov.
Efectos de serotonin
El personal de universidad de McMaster se hizo autores del trabajo científico durante el cual el gene clave de la obesidad - una especie de serotonin se ha encontrado. Es conocido que serotonin es �la hormona de alegría�, su desarrollo por un cerebro promueve la estabilidad emocional y el buen humor. Pero, como los investigadores canadienses explican, el serotonin que es responsable de emociones agradables pertenece al primer tipo de esta conexión.
�Se divide en dos tipos: en la escena de acción y en una forma de síntesis. El primer tipo se desarrolla en un cerebro e influye en emociones diferentes�, - los biólogos han explicado.
Al segundo tipo, serotonin periférico pertenece esta sustancia regula la actividad del tejido graso marrón del cual el desarrollo de la obesidad depende. Le también puede gustar Pankramin.
La grasa marrón contiene componentes que promueven la disminución en el azúcar en la sangre, y también a la incineración de calorías y su procesamiento en la energía. En un cuerpo de la persona hay ciertas zonas donde la grasa marrón se instala y sus jaulas son más activas, la cifra en la persona es más armoniosa. Los investigadores canadienses han venido a la conclusión que suprimiendo serotonin del segundo tipo; es posible aumentar la actividad bastante metabólica de células del tejido graso marrón. Por su parte, obligará un cuerpo �a quemar� la grasa blanca � y, ocurrirá independientemente de cómo la comida de la caloría alta es comida por la persona.
Receptores de Serotonin
Los efectos de serotonin son muy varios. Esta sustancia saques como un mediador en TsNS, influye en músculos lisos de buques y un GIT, participa en trombotsitarnom vascular un homeostasis. Los métodos de la reproducción molecular revelaron el de improviso gran número los receptores serotonin que basado en estructura y función se pueden dividir en 4 tipos. 5-T1--, 5-HT2-and 5-HT4-retseptory se integra a G-proteínas y a través de estas proteínas y los sistemas correspondientes de los segundos intermediarios influyen en funciones de varias enzimas y propiedades elek-trofiziologich de jaulas efectoras. Al contrario, 5-HT3-retseptory se atan con canales del ión. Aquí consideraremos stimulators y blockers los receptores serotonin. Las medicinas más nuevas de estos grupos que hacen funcionar selectivamente en subtipos separados los receptores serotonin se recibieron con trabajos con el uso de receptores recombinant. También nos pararemos en modelos experimentales que se aplican a una investigación de los medios que influyen en funciones mentales difíciles y su persistencia de violaciones, comportamiento agresivo, inquietud, depresión, desvelo del sueño del ciclo y otro. stimulators selectivos modernos de subtipos separados los receptores serotonin ya con el éxito se aplican en caso de migraña e inquietud y blockers selectivo en caso de varias violaciones gastrointestinales. Es posible influir en efectos fisiológicos de serotonin también por medio de los medios que actúan sobre la transferencia de serotoninergich. De este modo, los inhibidores de la captura de vuelta de serotonin eran medicinas eficaces para el tratamiento de depresión e inquietud.
A pesar de que el papel serotonin en muchos procesos fisiológicos y patológicos no levanta dudas, los puntos de su aplicación y los mecanismos de la acción se estudian mal. Quizás, tal situación es en parte causada por la variedad el serotoninovykh de receptores. Estos receptores grabados al principio por métodos farmacológicos son recibidos por KDNK que se reproduce hoy. Los receptores de Recombinant serotoninovy usan para estudiar de mecanismos moleculares del efecto de serotonin, y también para la búsqueda de los agentes que influyen selectivamente en subtipos separados de estos receptores. El círculo de un uso clínico de agentes similares se hace más amplio y más amplio.
Información histórica
En 1930mo Erspamer comenzó a estudiar la localización de células enterokhromaffin por medio de manchas en derivados de la índole. La concentración más alta de tales derivados fue grabada en el mucoso por un GIT; adelante había plaquetas y algunos departamentos de un CNS (Erspamer, 1966). Al ratito Peydzh y los coautores, que trabajan en la clínica de Clevelend, por primera vez emitieron la sustancia vasoconstrictive soltada de plaquetas en el curso de una parada de sangría y descifraron su estructura (Compenetración et al., 1948). Esta sustancia llamada por Paige un serotonin (Page, 1976), apareció a aquellos derivados de una índole que fue investigada por Erspamer. La descripción de caminos de la síntesis y la desintegración de un serotonin (Uden-amigo, 1959) y sus propiedades angiotonic (Sjoerdsma, 1959) permitió hacer una hipótesis según la cual las implicaciones de un llamado síndrome carcinoid en pacientes con tumores de células enterokhromaffin son causadas por la producción aumentada de esta sustancia. Realmente, tales pacientes tienen una eyección diaria con la orina de un serotonin y su metabolites puede alcanzar cientos de miligramos. Algunos síntomas de esta enfermedad hasta cierto punto indican mecanismos del efecto de serotonin. De este modo, en pacientes la psicosis similar al ácido que se levanta en la recepción se puede desarrollar. Al considerar que en animal y sustancias de tejidos de verduras con la acción alucinógena se encuentran similares con tripta-miny, es posible suponer que las sustancias similares se formen y causen una semiología psicótica en pacientes con el síndrome characinoid. Sobre la función del mediador de un serotonin en un cerebro de mamíferos se sugirió a mediados de años cincuenta (Brodie y Shore, 1957).
Los primeros datos de mecanismos moleculares del efecto de serotonin se obtuvieron en experiencias en un hígado de Fasciola hepatica (Mansour, 1979). Bajo la influencia de serotonin en ello la movilidad y la concentración de tsAMF bruscamente aumentaron; tanto esto, como el otro efectúa fueron bloqueados por un ácido. El aumento de la movilidad fue causado por phosphorylation tsAMF-dependiente de fosfofruktokinaza que limita glycolysis enzima. Sin embargo los receptores serotoninovy que median estos efectos en un hígado, probablemente, otros, que receptores de mamíferos conectados con adenilattsiklaz a serotonin. Como máximo tan datos detallados en mecanismos del efecto de serotonin no se sucedió para recibir hasta ahora.
Serotonin apareció en plantas y animales ya en etapas tempranas de la evolución, y a éstos, quizás, la abundancia de receptores serotonin habla (Peroutka y Howell, 1994). La reproducción de estos receptores mostró que algunas medicinas que se consideraron antes selectivas con relación a sus subtipos separados realmente tienen la afinidad alta con relación a varios subtipos. Más detalladamente sobre historia de estudiar y efectos de serotonin ver el artículo Sjoerdsma y Palfreyman (1990).
Propiedades químicas de serotonin
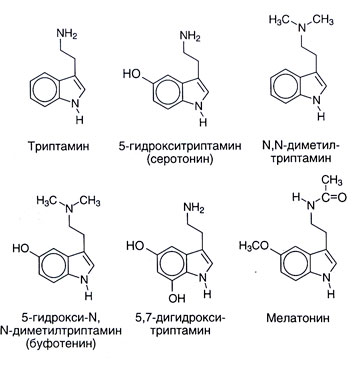
Fuentes. Dan la estructura química de serotonin y algunas conexiones cercanas con ello en fig. 11.1. Serotonin es extendido en una vida vegetal y animal: se encuentra en vertebrado, tunicates, moluscos, arthropods, celenterados, en la fruta y en nueces. También se encuentra en venenos � en una ortiga, en avispas y Escorpiones. Los numerosos parientes sintéticos o naturales a la sustancia serotonin también en un grado variado tienen los efectos fisiológicos centrales y periféricos. Muchos Ni O-metilirovannye indolamines (por ejemplo, N, N-dimetiltriptamin) son alucinógenos. Como se pueden desarrollar en un organismo, mucho tiempo los consideró posibles responsable de al menos algunas demostraciones de psicosis. Melatonin (5 metoksi N atsetiltriptamin) es formado de serotonin por N-acetylation con el O-methylation subsecuente (fig. 11.2). Esta sustancia sirve de indolamine principal del cuerpo donde su síntesis es regulada por factores externos (en particular, nivel de la iluminación). Melatonin causa depigmentation de melanotsit de la piel y suprime la función de ovarios. Quizás, desempeña un papel en biorritmos y por lo tanto puede ser útil en un síndrome de cambio de husos horarios.
Síntesis y catabolism. Un serotonin se forma del aminoácido irremplazable de un tryptophan en 2 etapas (fig. 11.2). En la primera etapa bajo la influencia de triptofangidroksilaza 5 hydroxytryptophan se forman, es la limitación serotonin reacción de síntesis. Triptofangidroksilaza representa un oxidase con las funciones mezcladas. El oxígeno molecular, y como la coenzima tetrahydrobiopterine participa en la reacción catalizada por ello. La actividad de triptofangidroksilaza, así como tyrosinehydroxylase, es regulada por phosphorylation, sin embargo el triptofangidroksilaza no es inhibido por el producto final en el mecanismo de feedback negativo. En un cerebro de triptofangidroksilaz no se satura con substrate y por lo tanto el precio de la síntesis de un serotonin depende de la concentración de un tryptophan. El último viene a células cerebrales por la captura activa por medio del transportista que es responsable del transporte de varios neutros y se bifurcó aminoácidos. En este aspecto el mantenimiento de un tryptophan en un cerebro depende no sólo de su concentración en el plasma, sino también en la concentración de otros aminoácidos que compiten con un tryptophan por un transportista.
Decarboxylation lleva 5 hydroxytryptophans a la formación de un serotonin. La disputa larga sobre si decarboxylases 5 hydroxytryptophans y DOFA son diferentes o la misma enzima, fue resuelta por KDNK métodos que se reproducen resultó que el mismo producto de genes es responsable de decarboxylation de ambos substrates. Ahora esta enzima se llama decarboxylase de L-aminoácidos aromáticos. Es muy extendido y afecta muchos substrates. 5 hydroxytryptophan son decarboxylized muy rápidamente y en un cerebro casi no se encuentra. En este aspecto las tentativas de afectar la concentración en un cerebro serotonin por la concentración cambian 5 hydroxytryptophans son condenados al fracaso.
El camino principal de catabolism serotonin la transformación en el ácido 5-hydroxyindolacetic que también sigue en 2 etapas (fig. 11.2). Al principio bajo la influencia de MAO el 5-hydro-xyindolacetaldehyde que entonces pasa en el ácido 5-gidroksi-indolacetic bajo la influencia de la enzima aldegiddegidrogenaza extendida en un organismo se forma (la cantidad insignificante 5 hydroxyindolacetaldehydes se convierten en el alcohol � 5-gidroksitriptofol). El ácido 5-hydroxyindolacetic es activly quitado de un cerebro; este proceso se suprime con un inhibidor no específico de probenetsidy de transferencia de transepithelial. Como en células nerviosas casi el 100% de todo metabolites de una caída de serotonin a la parte de ácido 5-hydroxyindolacetic, el precio de un recorrido de un serotonin en un cerebro se estima en la rebelión del nivel de ácido 5-hydroxyindolacetic después de la introducción de probenetsid. El ácido 5-hydroxyindolacetic que se forma en un cerebro y otros órganos, y también pequeñas cantidades de un 5-gidroksitriptofol y glucuronides se quita con la orina. Normal el diario egestion del ácido 5-hydroxyindolacetic en el adulto hace 2 � 10 mg. Valores más altos � un signo confiable de síndrome carcinoid. La síntesis bruscamente aumentada de un serotonin en esta enfermedad exige grandes números de pyridinic nucleotides y un tryptophan y por lo tanto los signos de la deficiencia de ácido nicotinic y un tryptophan no una rareza en tales pacientes. La rebelión de causas de etanol del mantenimiento de NADN, y a consecuencia de 5 hydroxyindolacetaldehyde pasa de un camino que se oxida de un catabolism a la recuperación (fig. 11.2).
Éstos son levanta egestion de un 5-gidroksitriptofol un poco y respectivamente reduce egestion de ácido 5-hydroxyindolacetic.
Hay dos isoenzymes de MAO: MAO A y MAO B. Al principio se dividieron basados en afinidad a substratos y sensibilidad frente a inhibidores; ahora tanto los isoenzymes se reproducen, como las propiedades de las formas reproducidas y naturales eran idénticas (Shih, 1991; también ver hl. 10). MAO A tiene la afinidad preferente a serotonin y noradrenaline, y su inhibidor selectivo es clorgilin. MAO B más trabajos de β-fenil-etilamin y benzylamine; MAO B inhibidor selectivo selegilin. La afinidad tanto de isoenzymes a dopamine como de triptamin es idéntica. Las jaulas nerviosas contienen tanto a MAO A como MAO B � principalmente en una membrana externa de mitochondrions. Ya que isoenzyme principal de plaquetas que también contienen serotonin en la alta concentración sirve a MAO B.
Se supuso que también hay otros caminos de un catabolism de serotonin, por ejemplo sulphation y Sobre - o N-methylation. El viaje final, en particular, podría llevar a la formación de una sustancia psychotropic endógena 5-hydroxy-N, N-dimetiltriptamina (bufotenina, fig. 11.1). Sin embargo otros metilirovanny indolamines (N, N-dimetiltriptamin, 5-metoksi-N, N-dimetiltriptamin) han mucho más expresado propiedades alucinógenas, y su papel en pathogenesis de psicosis es más probable.
El inactivation de serotonin es realizado no sólo por la desintegración enzymatic, sino también por medio de la captura de vuelta. Na + - el transportista dependiente localizado en una superficie externa de la membrana presynaptic de la terminación serotoninergich (proporciona el retiro de serotonin de una grieta sinóptica) y una superficie externa de una membrana de plaquetas es responsable de esta captura (extrae serotonin de la sangre). En plaquetas es el camino único de la acción del relleno de serotonin ya que no hay enzimas de la síntesis de esta sustancia en ellos. El transportista serotonin, así como otros transportistas de monoamines, se reproduce (hl. 12).
Puntos de aplicación serotonin
La mayor parte de todo el mantenimiento de serotonin en un organismo se cae a la parte de tejidos periféricos aunque también sirva de un mediador en un CNS. Su concentración en células enterokhromaffinnykh y plaquetas es la más alta. Un serotonin desempeña un papel importante en una regulación de un motility de un GIT.
Células de Enterokhromaffinny. Estas células calman en el mucoso un GIT. Sobre todo hay muchos de ellos en un duodeno. En las células enterokhromaffinnykh se sintetiza de un tryptophan y un serotonin se reúne, y también otros agentes biológicamente activos, por ejemplo la sustancia P y kinina contienen. Hay cierto nivel de la secreción básica de un serotonin en un GIT. Esta secreción amplifica en el estiramiento mecánico (por ejemplo, entrando en una nutrición o solución de sal hipertónica) y en una aburrida de fibras del motivo de nervios vagus. Quizás, el efecto estimulante de serotonin en un motility de un GIT también es mediado por su influencia en neuronas de plexus intermuscular (Gershon, 1991; también ver hl. 38). La secreción bruscamente aumentada de un serotonin y otros agentes biológicamente activos en el síndrome carcinoid es seguida de las perturbaciones gastrointestinales, cardiovasculares y nerviosas correspondientes. Además, la síntesis aumentada de un serotonin puede causar la deficiencia de ácido nicotinic y un tryptophan.
Plaquetas. Las plaquetas se diferencian de otros elementos formulados de una sangre, en particular, en la capacidad de tomar la tienda y soltar un serotonin. La síntesis de un serotonin en plaquetas no pasa. Un serotonin es plaquetas tomadas de una sangre y viene para el almacenaje al secreto elektronop gránulos por medio del transporte activo. Estos procesos son en muchos aspectos similares a captura y almacenaje de Noradrenalinum en las terminaciones comprensivas (hl. 6 y 12). A través de una membrana de plaquetas un serotonin es transferido por medio de Na + - transporte dependiente, y a gránulos � por el transporte activo secundario con el uso como una fuente de energía de un declive electroquímico para H + enmarcado por N +-Atfazoy. Al mismo tiempo la concentración de un serotonin en gránulos alcanza 0.6 mol/l, es 1000 veces más alto, que en cytoplasm de plaquetas. Precio En + - nuevo consumo serotonin dependiente por plaquetas un indicador sensible de actividad de inhibidores de nuevo consumo serotonin.
La función principal de plaquetas es hemostasis: cierran huecos en endothelium dañado. Por otra parte, la integridad de un endothelium desempeña un papel importante en el funcionamiento de plaquetas (Furchgott y Vanhoutte, 1989). El endothelium constantemente se pone en contacto a plaquetas como debido a las fuerzas de cambio que funcionan en la sangre corriente se desplazan a la periferia de buques (Gibbons y Dzau, 1994). El efecto de Vasoconstrictive de serotonin y tromboksan de A2 son resistidos por un factor endothelial de vasorelaxation (N0 y, quizás, algunas otras sustancias) (Furchgott y Vanhoutte, 1989; fig. 11.4). Para adherencia y agregación de plaquetas la condición de un endothelium tiene la importancia crucial (Hawiger, 1992; Ware y Heistad, L993). Cuando las plaquetas colindan con endothelium dañado, emiten las sustancias que causan su adherencia y liberación de un serotonin. ADF y ò ð îìáîêñàí A2 pertenecen a tales sustancias (hl. 26 y 55). Linkng de un serotonin con 5-HT2A-retseptorami tiene el efecto proagregant débil que amplifica bruscamente en la presencia de un collagen. Si el defecto de una pared vascular alcanza capas del músculo lisas, entonces un serotonin da el efecto vasoconstrictive directo que sirve de uno de mecanismos hemostasis. Este efecto amplifica bajo la influencia de agentes biológicamente activos que se emiten en el campo de daño tromboksan de A2, kinin, vasoactive péptidos. La formación de thrombi en un atherosclerosis es promovida por la destrucción de un endothelium y, como consiguiente, carencia de un factor endothelial de vasorelaxation. En estas condiciones la conducción de procesos a thrombogenesis siguen incontroladamente, como un círculo vicioso. Una parte en ellos también es jugada por un serotonin. El cuadro similar se puede observar en otras enfermedades de buques, por ejemplo el síndrome de Reynaud y vasospastic stenocardia.
La descripción a fig. 11.4. Funciones de un serotonin de plaquetas. La liberación de un serotonin de plaquetas es comenzada ellos por adherencia y agregación. Por su parte, un serotonin causa 1) la activación plaquetas de á-ÍÒ-receptors y, vrezultata, el cambio de una forma y aceleración de la agregación del último, 2) activación 5-HT, - los receptores similares de un endothelium con la asignación de un factor endothelial de vasorelaxation, 3) la activación de S-HT-receptors de músculos no estriados de buques y estrechamiento del último. Todos estos procesos siguen en la interacción con muchos otros agentes biológicamente activos y finalmente llevan a la sangría de la parada.
Sistema cardiovascular. La reacción típica de vasos sanguíneos a serotonin se estrecha. Los buques de órganos de GIT, riñones, pulmones y cerebro son sobre todo sensibles a ello. Serotonin también causa la reducción de músculos no estriados de bronquios. Sus efectos en el corazón son varios que el cambio de receptores serotonin de tonus de nervios vegetativos y reacciones reflejas es explicado por la activación de subtipos diferentes (Saxena y Villalon, 1990). De este modo, chronotropic positivo directo y el efecto inotropic de serotonin en el corazón pueden ser disfrazados por efectos de la exaltación de las fibras que van de receptores de la barra y chemoceptors. La influencia de un serotonin a las terminaciones afferent de nervios vagus causa Yarisha reflejo de Betsold quien muestran bradycardia agudo y la caída del ABP. A veces los arterioles bajo la influencia de un serotonin no se estrechan, y, al contrario, la supresión de la liberación de Noradrenalinum de las terminaciones comprensivas se extienden a consecuencia de la asignación de un factor endothelial de vasorelaxation y Prostaglandinums, y también. Por otra parte, un serotonin en sí mismo refuerza la acción vasoconstrictive de Noradrenalinum, angiotensin 11 e Histaminum. Promueve el efecto estático haemo aún más eficaz de serotonin (Gershon, 1991).
GIT. Probablemente, ya que la fuente principal y el almacenaje de un serotonin en un organismo sirven células enterokhromaffinny mucosas un GIT. El serotonin asignado por estas células atraviesa una vena portal a un hígado donde se metaboliza bajo la influencia de MAO A (Gillis, 1985). Alguna cantidad de serotonin pasa un metabolismo hepatic, pero rápidamente se toma un endothelium de tubos capilares pulmonares y también es afectada por MAO. El serotonin que se asigna en una pared de órganos de un GIT en su estiramiento mecánico o exaltación de nervios vagus participa en una regulación local de estos órganos. Bajo la influencia de un serotonin un motility de un estómago y un intestino puede tanto amplificar, y frenarse (Dhasmana et al., 1993) como en un GIT hay al menos 6 subtipos los receptores serotonin (etiqueta. 11.2). El efecto estimulante de serotonin es causado por su acción a las terminaciones de los nervios convenientes para capas musculares longitudinales y circulares (5-HT4-retseptory), en neuronas intramuros (5-HTj-and 5-NT|R-retseptory) y es directo en músculos lisos (5 receptores del IO en intestinos y 5-HT2B-retseptory en el día de un estómago). En un esófago el serotonin afecta 5-HT4-retseptory que en tipos diferentes de animales se puede seguir tanto reduciendo, como la relajación de músculos lisos. Los 5-HT3-retseptory (son muchos asistentes en las terminaciones de fibras sensibles del vagabundeo y otros nervios, y también en las jaulas) desempeñan un papel fundamental en un reflejo vomitivo (Grunberg y Hesketh, 1993). En una textura intermuscular las terminaciones serotoninergetic se encuentran. La liberación de serotonin en intestinos es causada por acetylcholine, la irritación de nervios comprensivos, y aumento de la presión intestinal intra y disminución en el pH (Gershon, 1991). El serotonin que se emite al mismo tiempo, por su parte, comienza reducir peristáltico.
El área principal de la concentración de cuerpos de neuronas serotonin en CNS es corazones de la costura de brainstem. Los procesos de estas neuronas van a todos los departamentos de una médula espinal y principal (hl. 12). Un serotonin se asigna no sólo en las terminaciones presynaptic, sino también en llamado varicosity de axons donde no hay exactamente synapses expresados (Descarries et al., 1990). En estos casos afecta muchas estructuras contiguas inmediatamente. Tal rasgo de la asignación y efecto de serotonin se compondrán con el punto de vista extendido que un serotonin no es sólo un mediador, sino también el modulador de la neurona (hl. 12).
En las terminaciones de neuronas serotonin hay todos componentes necesarios para la síntesis de un serotonin de un tryptophan (fig. 11.2). serotonin formado rápidamente viene a ampollas sinópticas donde en él MAO no puede trabajar. Después de la liberación en una hendidura sinóptica un serotonin otra vez se toma la terminación nerviosa por medio de Na - un transportista dependiente. Esta captura de vuelta sirve de un camino eficaz de un inactivation de un mediador. Las mismas moléculas de un serotonin que no vuelven a la terminación nerviosa son arruinadas por MAO localizado en neuronas postsynaptic y las siguientes células.
Efectos electrofisiológicos. Estos efectos de un serotonin se diferencian en áreas diferentes de un cerebro y en neuronas diferentes y dependen en que receptores afecta (etiqueta. 11.3; Aghajanian, 1995). Un serotonin puede tener la excitación y efecto del freno que discrepa en la dinámica temporal en las mismas neuronas. De este modo, en neuronas hippocampus un serotonin causa al principio la polarización hyper (causado por la activación de 1A-receptores 5-HT), entonces la despolarización lenta (causado por la activación de 5-HT4-retseptorov).
TsNS. Serotonin influye en TsNS muchas funciones, incluso un sueño, actividad cognoscitiva, percepción, dirección de movimientos, control termal, sensibilidad dolorosa, apetito, comportamiento sexual y regulación endocrina. En un cerebro todos se reprodujeron los receptores serotonin se encuentran, y a menudo en el mismo departamento hay varios receptores. Además, aunque la expresión de receptores serotonin en neuronas separadas se estudie insuficientemente, es posible creer que en la misma neurona varios subtipos de estos receptores se pueden localizar, y su activación se puede seguir tanto de synergistic como de efectos antagonistas. Puede ser la causa de una variedad extraordinaria de influencias de serotonin en funciones cerebrales.
Polarización de Hyper y disminución en la resistencia de una membrana que se levanta en caso de la activación 5-HT) A-receptores, son causados por el aumento de la permeabilidad del potasio. Estos efectos son bloqueados por la toxina, pero no dependen de tsAMF. Por lo tanto, pueden ser causados por el interfaz directo de 5-NT|A-retseptorov (a través de la G-proteína similar al soldado) con el canal del potasio (Andrade et al., 1986). La activación de los receptores localizados en un cuerpo y las dendritas de las neuronas de granos de una costura de un tronco de un cerebro también lleva a K + - polarización hyper dependiente. Aquí también participa en una emisión j de una señal de un receptor al canal sensible a la toxina de la G-proteína, pero el potasio corriente tiene otras características, que lo que se levanta en caso de la activación de postsynaptic 5-HT1A-retseptorov en un hippocampus. No se sabe aún por medio de que activación de mecanismos 5-HT1D-ayTopeuerrropoB lleva a la supresión de liberación de serotonin. Probablemente, el número de los canales calcic que se abren en respuesta a impulso nervioso las disminuciones potenciales.
La activación de 5-HT2A-retseptorov es seguida de la despolarización lenta. En algunos departamentos de un cerebro (por ejemplo, en la corteza prefrontal, un grano contiguo y un grano del motivo de un nervio facial) esta despolarización es causada por la disminución en la permeabilidad del potasio (Aghajanian et al., 1987). También hay otro mecanismo relacionado con la influencia en canales del ión; lleva al crecimiento de excitabilidad de la neurona y refuerzo de la reacción a mediadores emocionantes (por ejemplo, un glutamate). El papel del sistema fosfoinozitid en éstos efectúa aún no se establece. Cuando en la misma neurona también hay 5-HT1-retseptory y 5-Nt2d-retseptory, la reacción final a serotonin depende de una proporción entre la polarización hyper causada por la activación de 5-HT1-retseptorov y la despolarización causada por la activación de 5-HT2A-retseptorov. En el contexto de la polarización 5-Nt2d-blokatorov hyper amplifica. En muchas áreas de la corteza 5-HT2A-retseptory se localizan en las neuronas insertadas de Gamkergichesky y en neuronas piramidales. Por lo tanto, el estímulo de estos receptores puede llevar a influencias multidireccionales en neuronas piramidales en la dependencia porque si la acción en estas neuronas o en neuronas de Gamkergichesky prevalecerá. La activación de 5-Nt2s-retseptorov en oocytes de la rana, el expreso del m del ARN de estos receptores, lleva a la supresión del potasio corriente. En un cerebro tal efecto aún no se revela. El estímulo de 5-HT4-retseptorov que es seguido de la activación también causa la despolarización lenta causada por la disminución en la permeabilidad del potasio en neuronas. Aún no está claro por qué dos tipos diferentes de receptores serotonin conectados a varios sistemas de los segundos intermediarios dan el mismo efecto fisiológico. Además, la despolarización lenta es causada por la activación de uno más tipo de receptores serotonin 5 HT1P-receptores. Estos receptores se acompañan por el disponible sólo en neuronas intestinales intra y tienen propiedades farmacológicas especiales (Gershon, 1991).
La activación de 5-HT3-retseptorov causa la despolarización bystry. Se causa abriéndose del canal que pasa a Na + y K + (Higashi y Nishi, 1982) y hace un complejo uniforme (hemo canal sensible) con un receptor. La organización similar de un similar 5-HT3-retseptora a la organización N-holinoretseptora se ha confirmado en experiencias con la fijación local. Los 5-HT3-retseptory se encuentran en TsNS, ganglia comprensivo, fibras afferent paracomprensivas y comprensivas, intra neuronas intestinales y líneas celulares del origen neyronal (por ejemplo, NG108-15). En las propiedades farmacológicas 5-HT3-retseptory diferencian de otros serotonin receptores; quizás, hay varios subtipos de estos receptores que se diferencian en varias combinaciones de subjunits.
Funciones mentales. Los medios que actúan sobre receptores serotonin y varios cambios de funciones mentales. Muchos modelos experimentales queridos para una estimación provisional de la estimulación o bloqueo de la actividad de medicinas con relación a éstos o aquellos receptores, están basados en una investigación de tales acciones del motivo estereotípicas como, por ejemplo, reflejo. Las técnicas de Behaviouristic (por ejemplo, un método selecto de la medicina) permiten sugerir sobre un componente subjetivo de reacciones a medicinas psychotropic. Estas técnicas también se usan para una investigación de las medicinas que actúan sobre la transferencia de serotonin, en alucinógenos particulares (véase abajo). El análisis de una cantidad enorme de los trabajos dedicados a la influencia de serotonin en el comportamiento está más allá de nuestro libro, y sólo nos pararemos en aquellos experimentos que tienen una influencia directa a la psicopatología de la persona. Para el conocido más detallado con este sujeto es posible recomendar a las revisiones finas Glennon y Lucki (1988), Zifa y Fillion (1992), Koeketal. (1992).
Vaya en bicicleta un sueño es el desvelo. La regulación de un ciclo un sueño es el desvelo se hizo una de las primeras funciones mentales para las cuales el papel serotonin exactamente se estableció. Después del trabajo clásico de gatos que es realizado por Mouret y ñîò ð. (Mouret et al., 1967), el juego de datos que la reducción de la reserva serotonin por medio de causas el insomnio eliminado entrando de un precursor de serotonin 5 de aparecido. También pareció que tryptophan y no stimulators selectivo de receptores serotonin acortan el tiempo de dormirse y amplían la duración general de un sueño. Blockers de receptores serotonin se puede tanto levantar, y bajar una parte de un sueño lento profundo que, probablemente, es causado por la acción en subtipos diferentes de receptores serotonin (Wasquier y Dugovic, 1990). Tanto en animales como en blockers 5-HT2A-and 5-Nt2s-retseptorov (por ejemplo, riganserin) mejor dicho de fuentes fidedignas causan el aumento de una parte de un sueño lento profundo.
Agresión e impulsividad. Los datos obtenidos tanto en animales como en la persona, demuestre que un serotonin desempeña un papel importante en el comportamiento agresivo e impulsivo. En mucha comunicación de ensayos clínicos entre el nivel bajo de 5 - del ácido hydroxyindolacetic en SMZh y tal comportamiento se muestra (Brown y Linnoila, 1990). De este modo, la depresión de este nivel es ligada a tentativas impulsivas del suicidio (pero no con pensamientos suicidas; Virkkunen et al., 1995). Así como en caso de todos otros efectos de un serotonin, los animales no tienen datos finales de la influencia de un serotonin en el comportamiento agresivo aún aunque la asunción de tal influencia completamente se pruebe. Recientemente había apoyo de datos genético y la dilatación de tales visiones. Los receptores de Serotonin investigados por métodos de la ingeniería genética 5-HT) el V-receptor era el primero de. Por una nueva combinación homóloga la línea de ratones que tienen un gene que cifra este receptor se recibió era inactivated (Saudau et al., 1994). En tales animales la agresión más aguda se desarrolló lo que habla sobre un papel de estos receptores o en pasar con los contornos de neuronic que son responsables del comportamiento agresivo o es inmediato en la mayor parte de tal comportamiento. En la persona la mutación de punto del gene que cifra a MAO A se graba; la agresión al mismo tiempo extrema en la combinación con el retraso mental también se observa (Brunner etal., 1993). Como apareció, las implicaciones correspondientes también están disponibles para ratones con las mutaciones que causan la deficiencia de MAO A (Casos et al., 1995). Estos datos, seguramente, confirman una hipótesis de un papel de perturbaciones de la transferencia de serotoni-nergic en el comportamiento agresivo.
Inquietud y depresión. Eficacia de los medios que influyen serotoninergichesky transferencia (por ejemplo, inhibidores de la captura de vuelta de serotonin), en caso de frustración inquietante y depresión pruebas para ventaja de un papel serotonin en pathogenesis de estas condiciones. Sin embargo en modelos experimentales clásicos de éstos la frustración recibe resultados ambiguos el efecto de medicina depende de especies y clase de un animal y la técnica aplicada. De este modo, agonist parcial de 5-HT1A-retseptorov buspiron (hl. 19), siendo un tranquilizante eficaz en la persona, no reduce en la inquietud de animal en las experiencias entregadas por una técnica de preferencia o evitación; mientras tanto esta técnica se usó en caso del desarrollo de tranquilizantes. Al mismo tiempo el buspiron y otro blockers de 5-HT1A-retseptorov tienen el efecto anxiolytic completamente eficaz en otros modelos de la frustración inquietante (Barrett y Vanover, 1993). Los trabajos recientes de ratones con los genes inactivated de 5-NT1L-retseptorov también atestiguan para la ventaja de un papel de estos receptores en pathogenesis de la inquietud y, quizás, depresiones (Parques et al., 1998; Ramboz et al., 1998). Por otra parte, stimulators de unos los receptores serotonin, incluso 5-T2A--, inquietud de la causa de 5-HT2C-and 5-HT3-retseptorov tanto en animales de laboratorio, como en la persona. El papel de estos receptores y en modelos experimentales de una depresión se supone (por ejemplo, la impotencia entrenada).
La persona de datos directos de un papel serotonin en un pathogenesis de una depresión no tiene todavía bastante. Al mismo tiempo hay hechos clínicos muy brillantes. De este modo, en pacientes con unos efectos de la depresión de inhibidores de la vuelta serotonin nuevo consumo rápidamente se eliminan en las influencias que reducen serotonin nivel en un cerebro. Como tales influencias sirven, por ejemplo, la recepción de parachlorphenylalanine o las bebidas que no contienen un tryptophan, pero rico con aminoácidos neutros (Delgado etal., 1990). Es interesante que estas influencias en sí mismo no causen y no agraven una depresión. Significa que el nivel suficiente de un serotonin en un cerebro es necesario en primer lugar para la eficacia de inhibidores de su captura de vuelta.
Agentes que influyen serotonin nivel en tejidos
En los experimentos mandados en la aclaración de un papel fisiológico de un serotonin es posible aplicar blockers de receptores serotonin o los agentes que influyen serotonin nivel en tejidos. Hasta hace poco en la mayoría de trabajos sólo las medicinas del segundo de los grupos mencionados eran mecanismos usados de la acción de blockers los receptores serotonin se estudiaron mal.
En los gastos de explotación bajos de un tryptophan en una dieta serotonin nivel en un cerebro disminuye, en el contenido alto de aumentos de tryptophan. Como la reacción restrictiva de la síntesis de un serotonin se cataliza, los inhibidores de esta enzima causan la caída aguda del nivel de un serotonin. El más a menudo use un inhibidor selectivo irreversible de un triptofangidroksilaza parachlorphenylalanine. Bajo la influencia de esta sustancia serotonin nivel bastante y mucho tiempo disminuye, y el mantenimiento de catecholamins no cambia.
Parachloramphetamine y otras anfetaminas halogenated causan la emisión de serotonin de plaquetas y neuronas. En un cerebro después que allí viene la depresión larga de reservas de serotonin. Las anfetaminas de Halogenated son ampliamente usadas en trabajos experimentales. Dos de ellos fenfluramin y decsfenfluramin se aplicaron como anorecsant, sin embargo en relación a informes sobre su acción tóxica cardio en 1998 ellos en los EE. UU se retiraron de la venta. Las consecuencias del uso de estos agentes son hasta el final no estudiado. En las neuronas cerebrales serotonin causan expresado y largo (hasta varias semanas) depresión del nivel de un serotonin y el contenido de proteínas, específicas para estas neuronas que un transportista de un serotonin y triptofangidroksilaza al mismo tiempo disminuye. Puede demostrar la acción neurotoxin, sin embargo los signos de la muerte de neuronas bajo la influencia de anfetaminas halogenated no se encuentran. Derivados de triptamin con autoridades adicionales en lino de Indo un anillo (por ejemplo, 5.7-digidroksitriptamin; ver fig. 11.1) indudablemente causan la muerte las neuronas serotonin. La introducción de un 5.7-digidroksitriptamin un animal adulto es llevado a la destrucción selectiva por terminaciones serotonin, sin embargo los cuerpos de las neuronas no sufre, y con el tiempo las terminaciones se regeneran. Al contrario, en animales recién nacidos tanto las terminaciones como los cuerpos de las neuronas serotonin y por lo tanto el neogenesis no pasa fallecen.
Los inhibidores de la vuelta serotonin nuevo consumo, por ejemplo fluoxetine pertenecen a los agentes que influyen exactamente en la transferencia de serotonin. El mecanismo de su acción consiste en la extensión de efectos del serotonin asignado en la iniciación de las terminaciones nerviosas. Si junto con estas medicinas para entrar en 5 hydroxytryptophan, entonces las influencias de serotonin bruscamente amplifican. Los inhibidores de la vuelta serotonin nuevo consumo son uno de los antidepresivos más modernos y extendidos. El inhibidor de la vuelta serotonin nuevo consumo, Noradrenalinum y Dofaminum sibutramin se aplica como anoreksant. En un organismo dos metabolites activos que, probablemente, y tienen el efecto terapéutico se forman de él. Aún no está claro, influencia en que mediador causó el efecto de sibutramin.
Los inhibidores de MAO y Reserpinum pertenecen a no agentes selectivos que influyen serotonin nivel en tejidos. Los inhibidores de MAO bloquean el camino principal de un metabolismo de un serotonin, y Reserpinum causa su emisión del almacén de la neurona con el desgaste subsecuente. Todos estos agentes llevan a la depresión expresada del mantenimiento de serotonin, sin embargo al mismo tiempo en el mismo grado también el nivel de disminuciones de catecholamine. Por lo tanto ya que los reactivos para los inhibidores de análisis farmacológicos de MAO y Reserpinum se usan rara vez. Se aplicaron en la psiquiatría: Reserpinum como narcoleptic, e inhibidores MAO como antidepresivos.